Part A
Purpose:
This segment of the lab is designed to familiarize the experimenter with the concept of phases of cellular mitosis in both plant and animal cells, as wells as the duration that each lasts for. On the whitefish and root tip slides, all four phases of mitosis exist, in varying numbers. To determine which stages are longest, one needs only count the cells and identify the stages they are in. The more of that stage existing at one time on the slide, the longer the stage.
Introduction:
Mitosis is the process that cells go through in order to divide. There are four encompassing terms to divide this process into easy to understand steps. The first is prophase, the point in time where chromatin condenses into chromosomes in order to facilitate the dividing process and equally split genetic information amongst daughter cells. At this point in time, the cell is undergoing basic changes to prepare itself for division. The second is metaphase, the point in time at which the cells’ chromosomes align in the center and attach to the spindle from their centromere. At this stage, the cell is preparing to divide and is separating the chromosomes as such. The third is anaphase, where the chromatid pairs separate and walk along spindle fibers towards opposite sides of the cell. At this point in time, the cell is almost ready to divide since the genetic information is (usually) evenly distributed. The final phase is telophase, where a new nuclear envelope envelopes the chromosomes, marking the beginning of nucleus formation. After which, the process of cytokinesis splits the cell in two and creates two identical daughter cells. In plant cells, a new cell wall is formed in between the new cells rather than the animal cell’s cleavage furrow process.
Methods:
Two partially separate, but equally interconnected tests were undertaken in order to quantify the length of time cells spend in each stage of mitosis. The first step was actually discovering which cells were in which stage. Using a microscope on the 10x objective setting, a location on both the whitefish and root tip was focused on, and the 40x setting was used to study individual cells. From this process, and prior knowledge, how each cell looked in each stage was made apparent.
Interphase

Whitefish
This example boasts an intact nuclear envelope, which allows us to infer that cell is not ready to divide and is in the process of duplicating genetic information, organelles, and growing in size.
Prophase
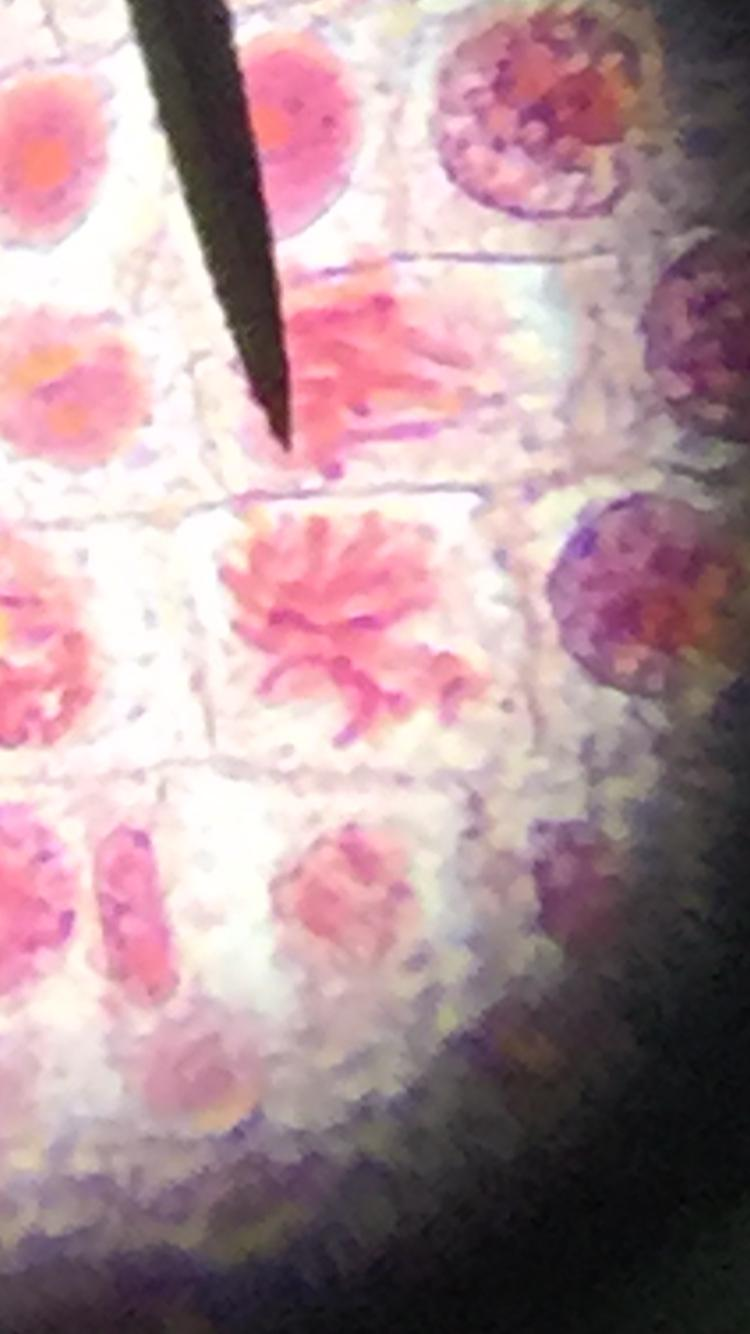
Root Tip
This image shows that chromosomes have begun to form in this phase, but are unorganized.
Metaphase

Root Tip
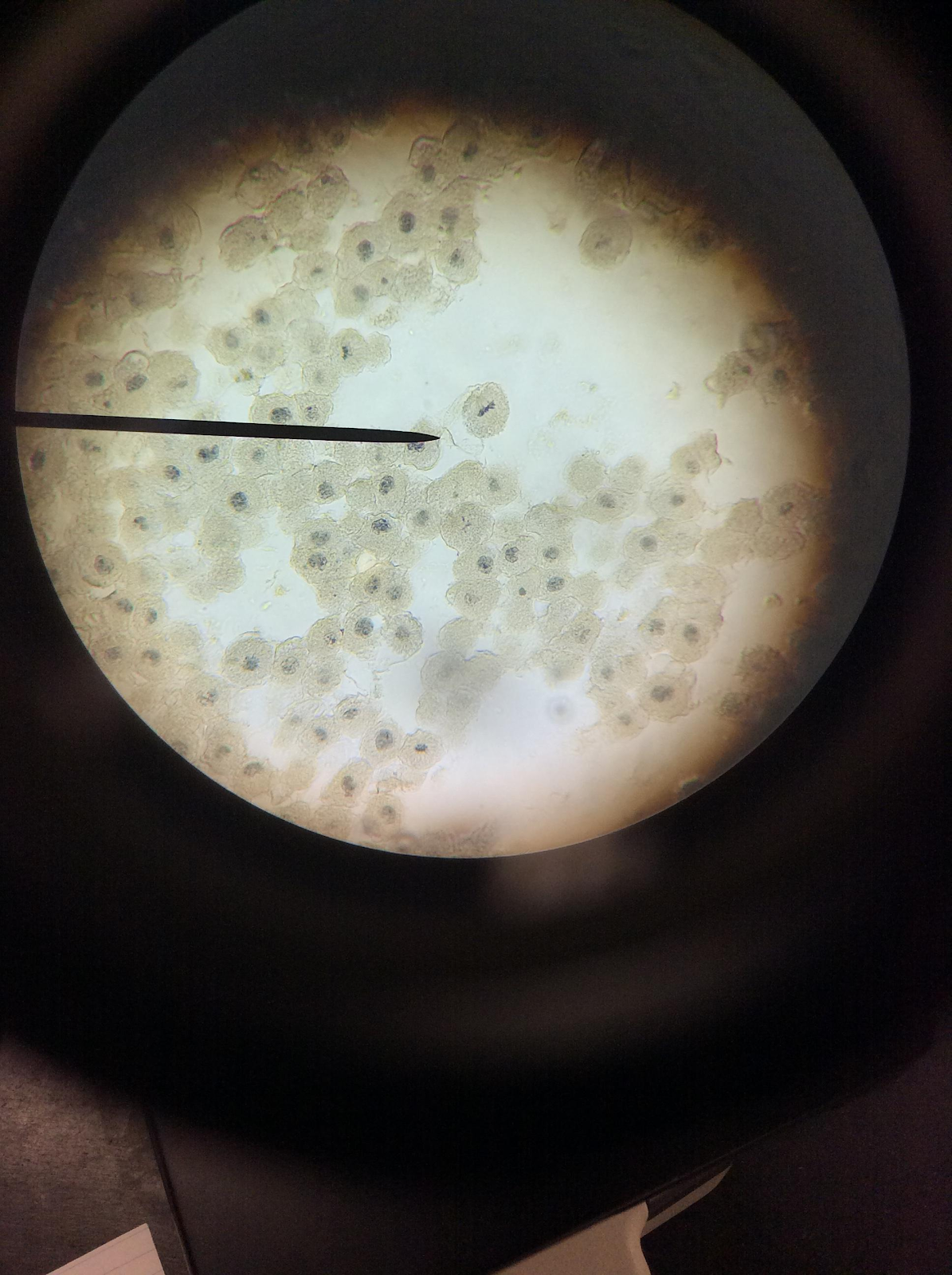
Whitefish
These two images show how chromosomes begin to line up along the center of the cell during this phase.
Anaphase

Root Tip
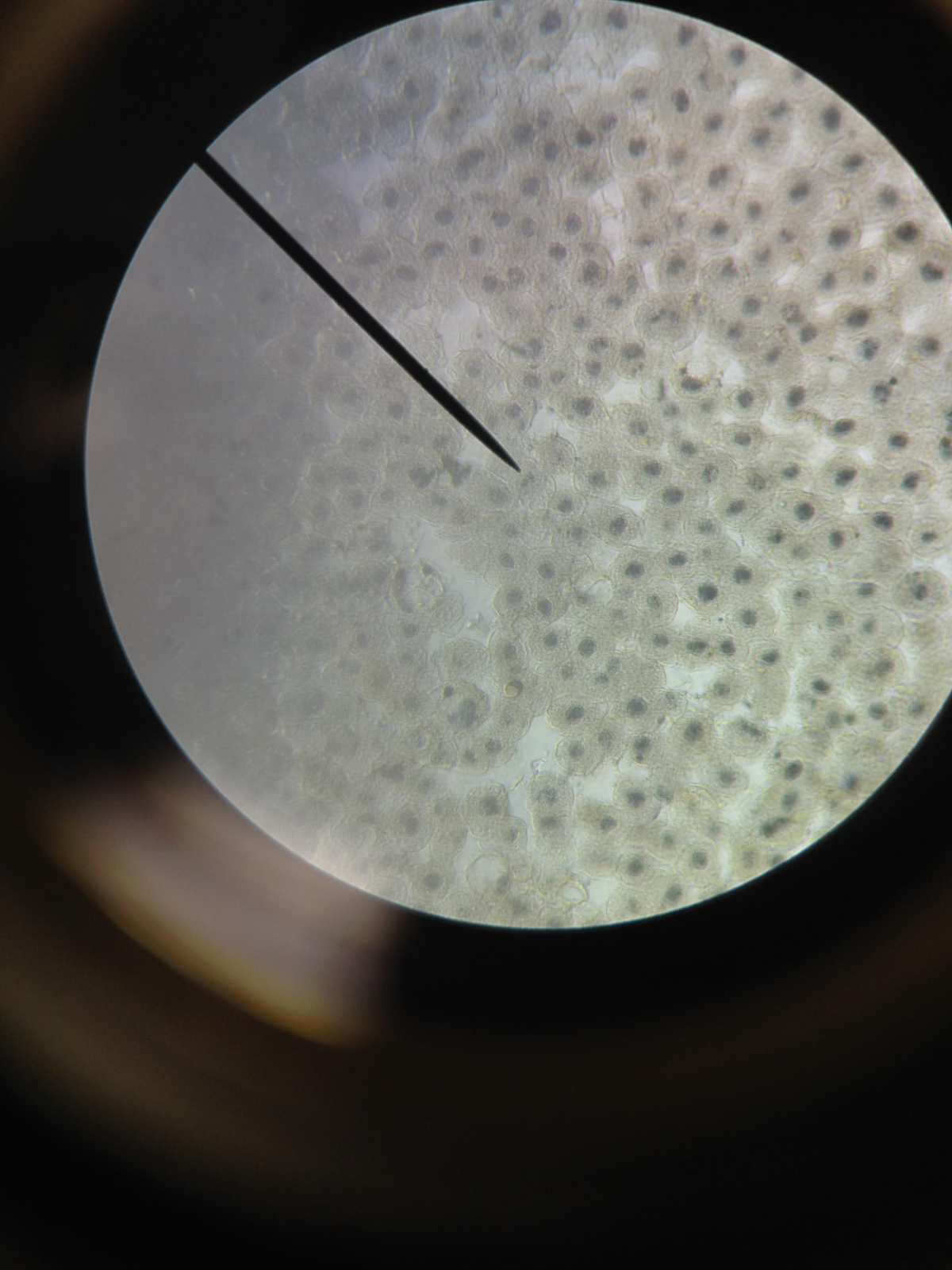
Whitefish
These two examples (pardon the image scaling on the whitefish) demonstrate how chromosomes walk along their spindle fibers during this phase.
Telophase
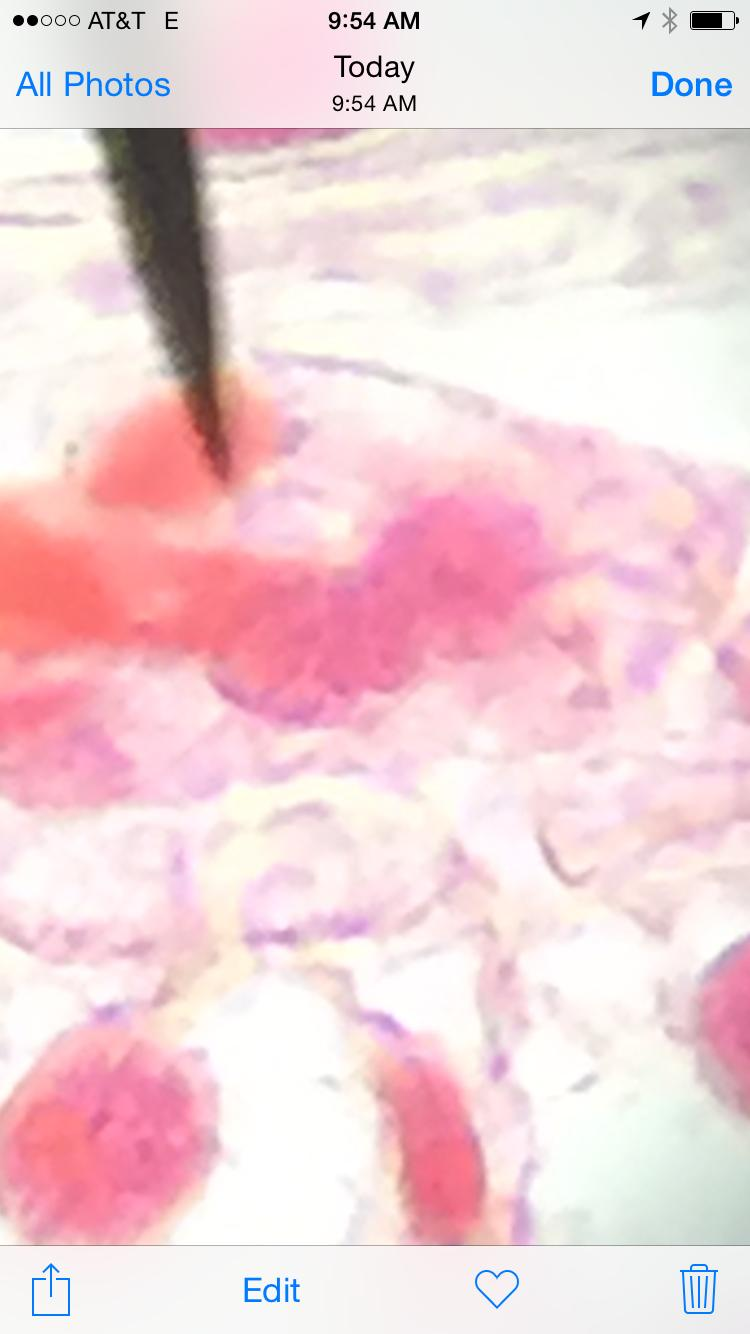
Root Tip

Whitefish
These two examples show how the cell splits in two during this phase to create two identical daughter cells.
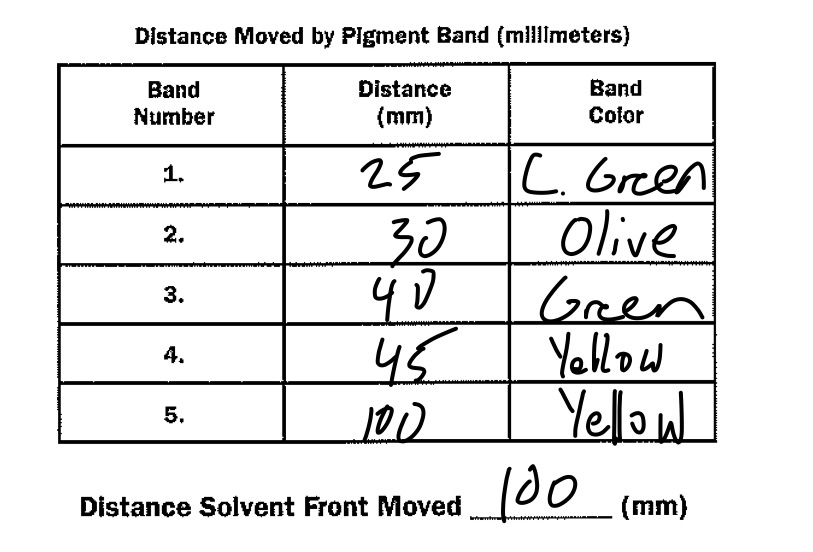
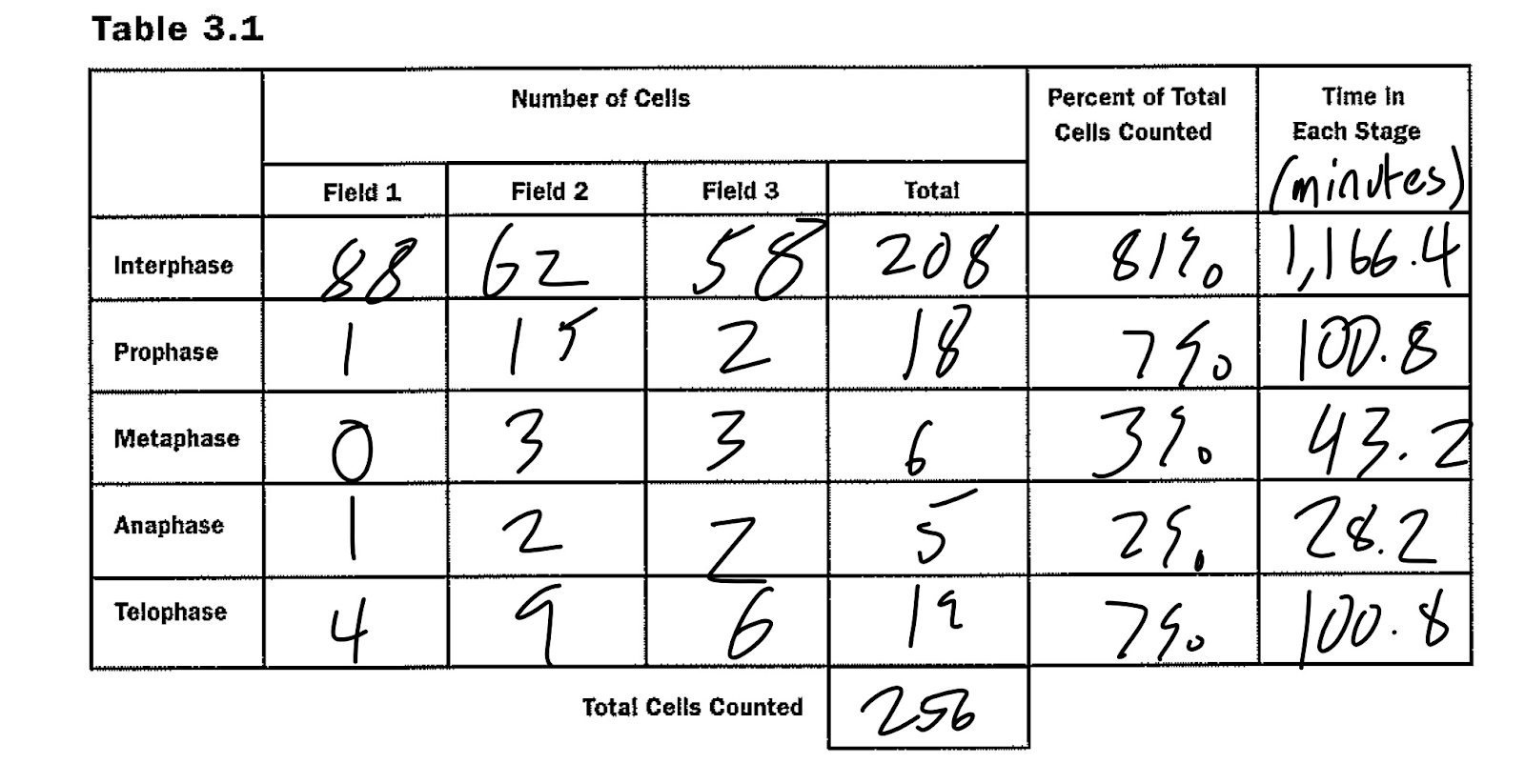
With this knowledge in hand, we were able to quantify how many cells were in each particular stage of mitosis by using an extraordinarily simple process: counting! If the cell appeared to look like the example, it was tallied under the category related to the specific stage of mitosis. This counting process took place on three different fields, and the total number of cells and what stages they were in were counted for each. Once that data was retrieved, the total number of cells in each stage was added together, and the percentages were calculated based against the total number of cells. Afterwards, this percentage was multiplied by 1440 minutes in order to generate the time a standard cell spent in that particular stage. The results from this can be viewed in the below table.
Data:
This chart organizes the data collected in the latter part of the procedure.
Graphs and Charts:
No graphs were made in this particular experiment.
Discussion:
During interphase, the cell is preparing to divide. It does this by growing much larger, duplicating its genetic information (evident by the fact that there are two sets of chromosomes) and duplicating its organelles in order to facilitate proper growth and function of the daughters. When the cell duplicates, it has all the basic properties it needs to survive. All it needs to do is grow and duplicate its own genetic information and organelles so it, too, can divide.
In plant cells, mitosis works nearly identically to animal cells. However, instead of pinching apart, a new cell wall is formed to separate the daughter cells. This is because the plant’s cell wall is not a flexible membrane, and does not allow the cell to pinch apart. This rigid structure is what gives plants their characteristic rigid appearance.
Centrosomes are parts of the nuclear membrane that break off during prophase and form the mitotic spindle, which allows the chromosomes to walk across to opposite sides of the cell during later phases. If these centrosomes did not exist, cellular division could not occur since chromosomes would be unable to migrate themselves to the future daughter cells.
In regards to the specific root tip we observed, it was actively dividing. Many cells were in the mitosis stage, but in separate phases. If this locale was not actively dividing, even more cells than there currently are would be in interphase instead.Since mitosis only lasts a few hours whereas interphase can last around 18-20, the number of cells in a specific mitosis stage would be drastically reduced to a number even smaller than this experiment boasts now.
In regards to the table, we can assume that cells spend the most time in interphase. This makes sense, as the cell needs a lot of time to literally duplicate its interior components, like organelles and genetic information, as well as growing to a size that would be able to produce daughter cells large enough to sustain themselves. The next stages are tied between prophase and telophase. The former condenses chromatin into chromosomes, which then migrate to more organized formations in later stages, whereas the latter splits the cell in two after forming a nuclear envelope around the chromosomes. This can lead us to believe that the formation or degradation of chromosomes into or from chromatin is a time-consuming process. Considering that the matter is essentially changing states from a viscous liquid to a more solid counterpart, this change makes sense. Metaphase is third longest, which entails the process of organizing chromosomes at the center of the cell and attaching them to spindle fibers. From this, we can infer that this organization is a time-consuming process, most likely to ensure that DNA is separated equally in the following phase to prevent a cell from having an improper amount of chromosomes. And the shortest stage is anaphase, which is solely the process in which the chromosomes walk along the spindle fibers. This is the only event that happens in this stage, making it understandable why it takes the least amount of time.
Conclusion:
This portion of the lab was designed to ensure that the experimenter understands why mitosis and interphase take the amount of time that they do. Interphase is the longest because the formation, duplication, and growth of important cellular features are vital to the state of the cell post-division. Mitosis is comparatively shorter, but the stages it is divided into are all unique and paramount to the formation of a functioning cell. If anything goes wrong, a cell can mutate. It could be a lack of a certain chromosome or organelle, but many mutations are a detriment to the organism as a whole. Cancerous cells grow without regulation, forming tumors that can kill organisms quickly if they spread. Certain mutation in gametes (it’s meiosis, but the same idea applies) can cause mental conditions that stunt the ability for an organism to function. In short, each stage of the mitosis process is important and paramount to the organism’s survival.
References:
- Unless otherwise stated, all information related to mitosis has been synthesized from Jane Reece’s Campbell Biology AP Edition Textbook and Collegeboard’s Lab Three: Mitosis and Meiosis.
- Tables 3.1’s outline has been taken from Collegeboard’s Lab Lab Three: Mitosis and Meiosis, but the information is original to the experiment.
- Paraphrased procedures have been synthesized from procedures found in Collegeboard’s Lab Three: Mitosis and Meiosis
- All pictures are original from the in-class experiment.
Part B
Purpose: In this lab we wanted to show the different stages of meiosis using chromosome models. The models were a representation of a chromosome going through both Meiosis I and Meiosis II. Nonetheless, it was used to show how crossing over occurs in genes.
We used these models to observe meiosis I and meiosis II at the chromosomal level at a size that was easier to understand.
We used these models to observe meiosis I and meiosis II at the chromosomal level at a size that was easier to understand.
Introduction:
Meiosis produces four unique haploid cells unlike mitosis which produces two identical diploid cells. In Meiosis I, the chromosome number changes from diploid to haploid. Moreover, in Meiosis II, the sister chromatids are separated which creates four haploid cells. Crossing over increases genetic diversity it occurs when two different chromosomes exchange their genetic material. The distance between two chromatids determines how much crossing over occurs. Having more space allows crossing over to occur more frequently .
Meiosis produces four unique haploid cells unlike mitosis which produces two identical diploid cells. In Meiosis I, the chromosome number changes from diploid to haploid. Moreover, in Meiosis II, the sister chromatids are separated which creates four haploid cells. Crossing over increases genetic diversity it occurs when two different chromosomes exchange their genetic material. The distance between two chromatids determines how much crossing over occurs. Having more space allows crossing over to occur more frequently .
Methods:

Interphase - in Interphase, chromosomes start as single chromatin strands that are not organized. Then, DNA synthesis occurs, joining the single strand to a sister chromatid at the centromere. This creates a chromosome.

Prophase I - In this phase, the two homologous chromosomes pair up to exchange genetic information via crossing over as well as form a tetrad. This is different than prophase in mitosis where the nuclear envelope begins to disintegrate. The crossing over of chromosomes that is shown is the primary reason for genetic variation among species.
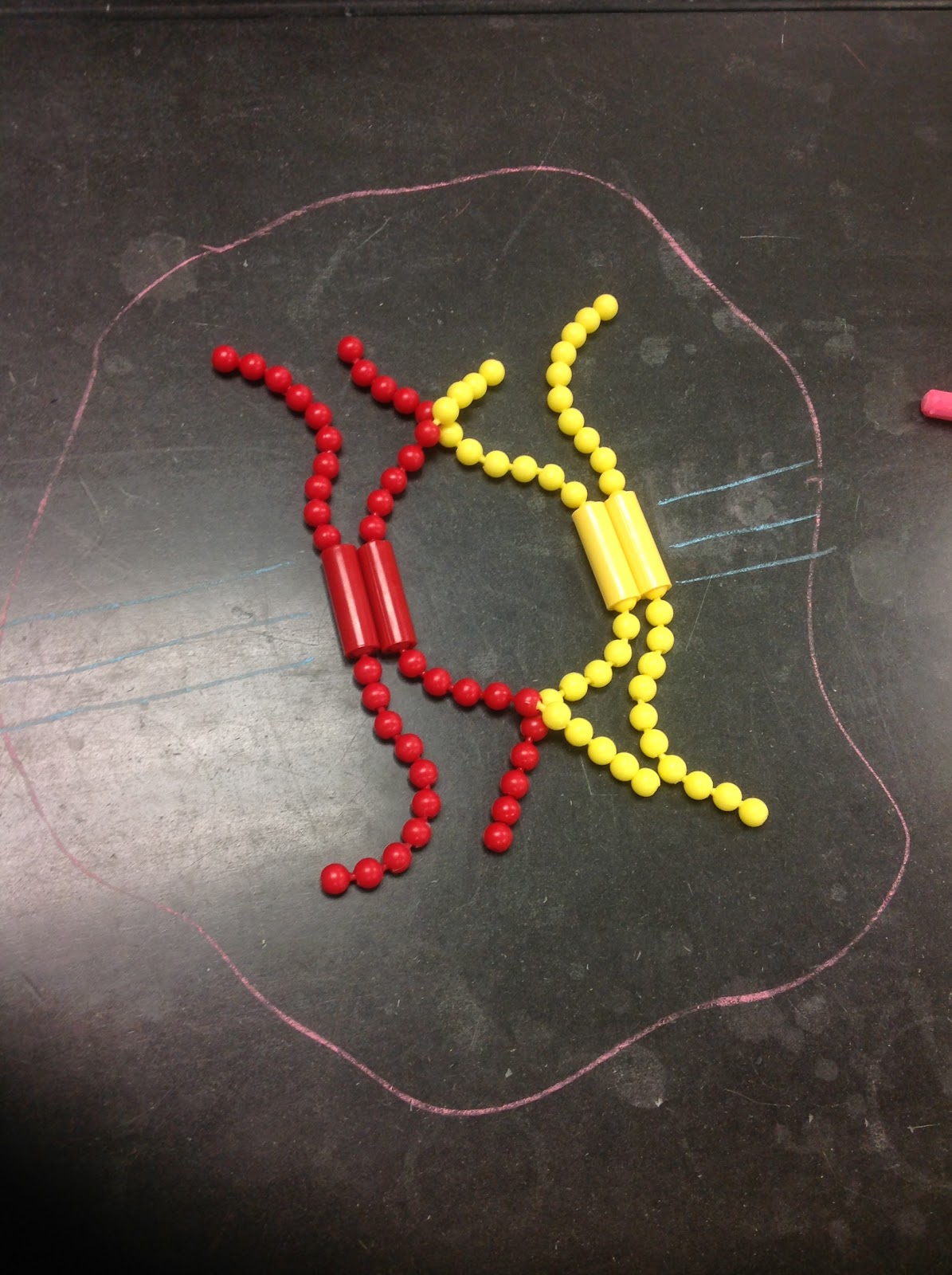
metaphase I - The newly formed tetrad said move to the center of the cell, between the two poles along the “metaphase plate”. In Metaphase I, the spindle fibers attach the centromeres of each chromosome. Unlike Metaphase in Mitosis, the kinetochores of the sister chromatids face the same pole.

Anaphase I - in this phase, the homologous chromosomes are separated and pulled to opposite poles along the spindle fibers. This differs from mitosis, where the sister chromatids are torn apart at the centromere and pulled to opposite poles.

Telophase I - In this phase, a nuclear envelope forms, as well as a cleavage furrow. Then, two haploid cells are produced, both with chromosomes that still have both sister chromatids.
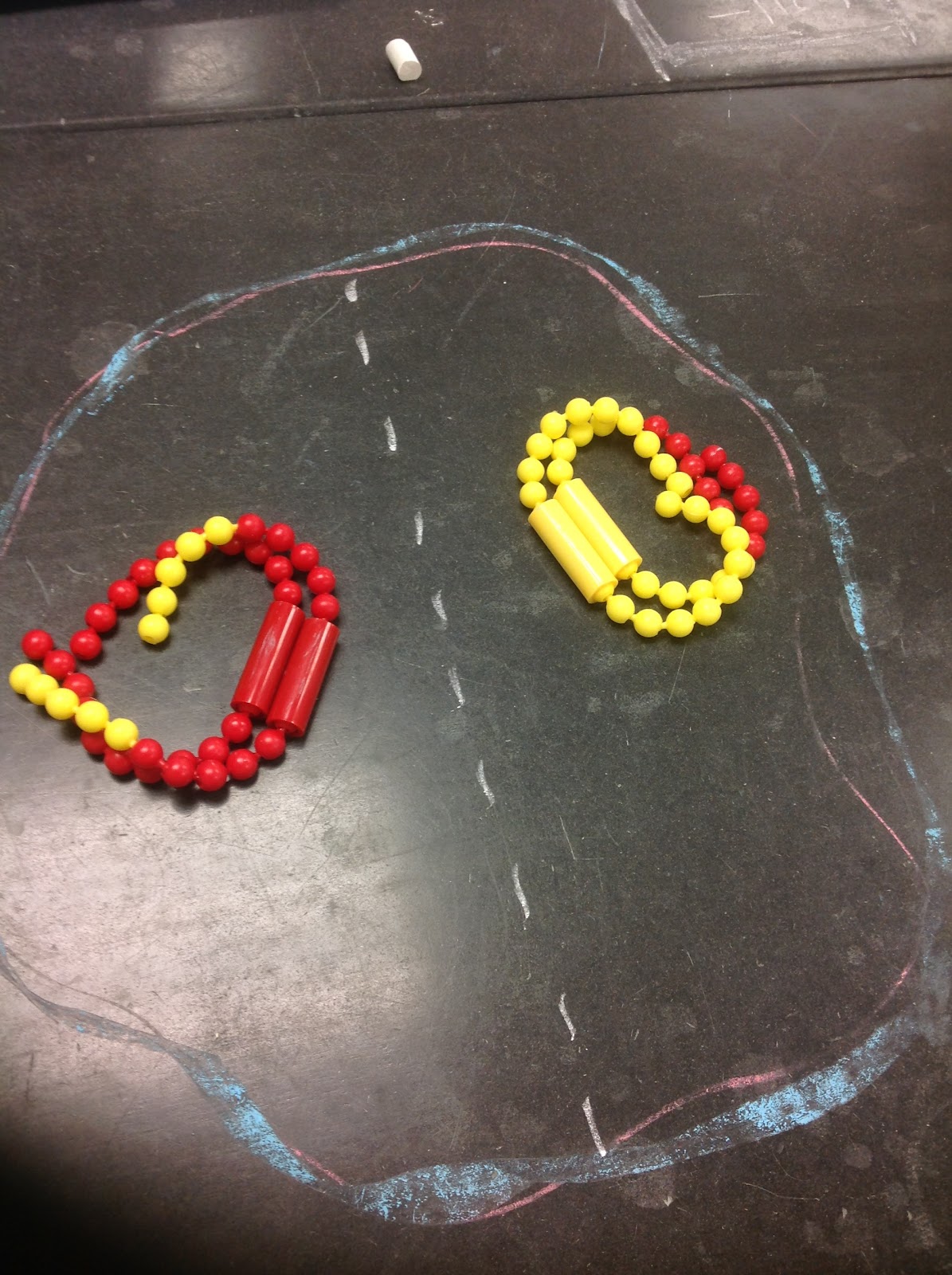
Prophase II - In this phase, we start off with the two haploid cells we left off with in telophase I. In oogenesis and spermatogenesis, these cells are the secondary oocytes and spermatocytes respectively. The nuclear envelope begins to break down and the centromeres move towards the poles.

Metaphase II - Again, the chromosomes line up in the middle of their respective cells along the metaphase plate. Then, similar to mitosis, the spindle attaches the kinetochores of each centromere to the opposite poles. This results in the separation of sister chromatids later on in Anaphase II.

Anaphase II - the sister chromatids are pulled apart by the spindle fibers towards opposite poles of each cell. They are now chromosomes. The cells begin to elongate in preparation for cytokinesis to occur.

Telophase II - The final phase of meiosis, the chromosomes arrive at opposite sides of the cell. Cytokinesis occurs, forming genetically varying haploid cells. In oogenesis, one of these cells is a mature ovum, while the rest are polar bodies to be reabsorbed by the body. In spermatogenesis, all four haploid cells are spermatocytes to be used in sexual reproduction.
Data:

Discussion:
There are many stages of meiosis I and meiosis II. In the first stage of meiosis I, interphase I, chromatin is found coiled inside the nuclear envelope. Karyokinesis, the division of the nucleus of a cell is a process that occurs during interphase I. The nucleus must divide before the cell can divide. The centrosomes that are located within the plasma membrane are in close contact and yet they do not form spindles. In the next stage, prophase I, spindles begin to be formed within the plasma membrane. In metaphase I the centrosomes begin to line up where they attach to the kinetochores. The mitotic spindle continues to enlarge during this phase of meiosis I. In meiosis I a cell is able to divide into two diploid cells; however, in meiosis II those two diploid cells divide into four haploid cells. Meiosis plays a crucial role in sexual reproduction because it produces haploid sex cells which contain the right amount of chromosomes. Through meiosis greater genetic diversity is achieved which is beneficial because it gives organisms a greater chance to survive.



Conclusion: From our data we found that mitosis and meiosis have very different end results. For example, in meiosis the number of chromosomes are cut in half and four daughter cells are produced. Mitosis on the other hand produces two identical cells. We observed that meiosis I differs from mitosis in almost every phase, most noticeably in anaphase I and telophase I where the homologs are separated (As opposed to sister chromatids in mitosis) and the resulting cells are haploid (As opposed to Diploid in mitosis). Then, we were able to observe the similarities between meiosis II and mitosis, with the only major differences being that the cells in meiosis II are haploid and the resulting number of cells is four. Meiosis II acts out very similar to Mitosis, with spindle fibers of opposite poles attaching to kinetochores in Metaphase II, and the resulting separation of sister chromatids in Anaphase II causes there to be 4 gametes created after telophase II because the chromosomes from each cell are still attached to their sister chromatids.